Снижение содержания афлатоксина М1 в молоке с помощью куркумина: кинетическое моделирование методом Монте-Карло (исследование in silico)
Снижение содержания афлатоксина М1 в молоке с помощью куркумина: кинетическое моделирование методом Монте-Карло (исследование in silico)

Аннотация
Афлатоксин M1 (AFM1), это термостабильный канцерогенный микотоксин, выделяемый с молоком лактирующих животных после употребления кормов, загрязнённых афлатоксином B1. Глобальная распространённость этого токсина в молочных продуктах во многих регионах продолжает превышать установленные нормативы. Куркумин, липофильный полифенол из куркумы длинной (Curcuma longa L.), предлагается в качестве природного деконтаминанта; однако конкурентное связывание куркумина молочными белками может существенно снижать его эффективную свободную концентрацию, что приводит к систематическому завышению эффективности деконтаминации в простых матричных моделях. Была разработана вычислительная структура на основе моделирования Монте-Карло (1000 итераций) для кинетического описания снижения концентрации AFM1 в течение 24 часов при дозах куркумина 0; 2,5; 5 и 10 мг/л. Сравнивались две механистические модели: модель линейного распада первого порядка (Модель А) и модель насыщения Михаэлиса–Ментен с учётом конкурентной интерференции молочных белков (Модель В). Для всех параметров были заданы априорные логнормальные распределения, что соответствует правосторонней асимметрии данных о встречаемости микотоксинов. Для разделения предсказательной дисперсии между параметрами модели использовали анализ чувствительности Соболя. Модель насыщения выявила явное плато эффективности при более высоких дозах куркумина, которое не фиксировалось линейной моделью. При дозе 2,5 мг/л Модель В достигла более высокого соответствия нормативам (98%), чем Модель А (85%), благодаря более благоприятной кинетике при дозах ниже насыщающих; при дозах 5 и 10 мг/л обе модели предсказывали практически полное соответствие максимально допустимому уровню (МДУ) ЕС, составляющему 0,05 мкг/кг. Максимальная скорость удаления (Vmax; индекс Соболя Si = 0,697) была идентифицирована как подавляющий источник предсказательной неопределённости, за которым следуют начальная концентрация AFM1 (Si = 0,160) и константа половинного насыщения (Si = 0,135). По сведениям автора, в данном исследовании впервые представлена вероятностная кинетическая структура, количественно определяющая влияние интерференции молочных белков на снижение AFM1, опосредованное куркумином, что даёт практический инструмент для риск-анализа с учётом неопределённости и априорного планирования экспериментов в молочных системах.
1. Introduction
Aflatoxin M1 is the main hydroxylated metabolite of aflatoxin B1, formed in the liver of lactating animals following ingestion of contaminated feed and subsequently excreted in milk. Owing to its thermal stability and resistance to standard pasteurisation conditions , , aflatoxin M1 poses a well-known carcinogenic risk in commercial dairy products . The European Union (EU) enforces an MRL of 0.05 µg/kg for AFM1 in raw and heat-treated milk . A recent worldwide systematic review and meta-analysis reported a mean AFM1 concentration of 57.36 ng/L in raw milk, highlighting the need for continuous monitoring and effective control strategies . Despite this evidence, several surveys continue to report cases of AFM1 contamination exceeding the MRL in multiple regions .
Current mitigation strategies for AFM1 in milk focus on physical adsorption using bentonite or activated carbon, and biological binding using lactic acid bacteria and yeast cell walls , , . While effective under experimental conditions, these approaches can alter the organoleptic properties of milk or encounter consumer acceptance barriers. Growing interest in “green chemistry” decontamination using plant-derived polyphenols has directed attention to curcumin, the principal bioactive polyphenol of Curcuma longa L. . Curcumin is a lipophilic compound capable of interacting with hydrophobic xenobiotics via π–π stacking and hydrogen bonding , . Its application as a direct milk additive has been explored in the context of antioxidant properties and safety , , yet its kinetic capacity to reduce AFM1 concentrations under realistic dairy matrix conditions has not been quantitatively characterised. Curcumin is not organoleptically neutral either: at concentrations such as 2.5–10 mg/L, it imparts a distinct yellow colour and a mild earthy, peppery flavour. However, curcumin is already approved as a food colourant (E100) and is used in certain dairy products (e.g., yellow cheeses, spiced fermented milks). Therefore, its sensory acceptability depends on the target product and consumer expectations
.An important methodological gap in the existing literature is the absence of kinetic modelling frameworks that study parameter uncertainty. Most experimental studies report only static percentage removal values, which do not capture the dynamic nature of toxin–binder interactions , . Moreover, milk is a complex colloidal system in which caseins and whey proteins bind polyphenols with high affinity , , reducing the free fraction of curcumin available for interaction with AFM1. This competitive sequestration implies that AFM1 removal kinetics in milk are likely non-linear and saturable rather than strictly first-order. A probabilistic Monte Carlo simulation framework, drawing on methodology applied to mycotoxin mitigation research , offers a means of quantifying not only central tendency estimates but also the probability of achieving regulatory compliance under conditions of parameter uncertainty. The present study applies this approach to compare a standard first-order decay model against a saturable Michaelis–Menten model, to provide a quantitative, uncertainty-aware basis for evaluating curcumin as a potential hurdle technology for AFM1 mitigation in milk.
2. Research methods and principles
2.1. Study Design
This work is a theoretical in silico investigation. No biological samples were processed, and no laboratory experiments were conducted. The simulation study models the kinetic trajectory of AFM1 concentration C(t) in liquid whole milk over 24 hours under varying curcumin doses. Two competing kinetic formulations are compared to evaluate the effect of milk matrix interference on predicted decontamination efficacy.
2.2. Parameter Distribution
The initial AFM1 concentration (C₀) was sampled from a log-normal distribution with arithmetic mean 0.08 µg/L and arithmetic standard deviation (SD) 0.03, reflecting the right-skewed character of mycotoxin contamination survey data and representing a plausible contamination scenario above the EU MRL. Curcumin doses were set at 0, 2.5, 5, and 10 mg/L, consistent with reported solubility limits in colloidal dairy systems . All kinetic parameters were assigned log-normal prior distributions (see Table 1). Parameter ranges were derived by analogy from literature on polyphenol–protein binding interactions and microbial adsorption isotherms in dairy matrices , , as curcumin-specific kinetic constants for AFM1 interaction in whole milk have not yet been experimentally determined. The dose-specific potency coefficient α (Model A) was treated as a fixed constant of 0.010 h⁻¹·(mg/L)⁻¹ .
Table 1 - Input parameters and probability distributions used in the Monte Carlo simulation
Note: AFM1 = aflatoxin M1; SD = standard deviation (arithmetic). Vmax and Km are specific to Model B. The potency coefficient α = 0.010 h-1·(mg/L)-1 is a fixed constant for Model A and is not subject to Monte Carlo sampling
Parameter | Symbol | Distribution | Mean | SD |
Initial AFM1 concentration | C0 | Log-normal | 0.08 µg/L | 0.03 |
Natural decay rate constant | k0 | Log-normal | 0.005 h⁻¹ | 0.001 |
Maximum removal rate (Model B only) | Vmax | Log-normal | 0.12 h⁻¹ | 0.03 |
Half-saturation constant (Model B only) | Km | Log-normal | 3.0 mg/L | 1.0 |
2.3. Kinetic Models
Model A (first-order linear decay) assumes AFM1 removal is directly proportional to curcumin dose, without capacity limitation:
where k0 is the natural degradation rate constant (h-1), α is the dose-specific potency coefficient (h-1·(mg/L)-1), and D is the curcumin dose (mg/L).
Model B (Michaelis–Menten saturation) introduces a saturation term representing competitive sequestration of curcumin by milk matrix proteins:
Here, Vmax (h-1) is the maximum achievable removal rate, and Km (mg/L) is the dose required for half-maximal removal efficiency, reflecting both the intrinsic affinity of curcumin for AFM1 and the apparent attenuation of curcumin availability due to competitive protein binding in the milk matrix.
2.4. Statistical Analyses
All simulations were performed in R (v. 4.3.2; R Core Team) . Monte Carlo sampling comprised 1000 iterations per dose level, yielding 4000 kinetic trajectories in total. Sobol first-order sensitivity indices were computed using the sensitivity package in R to partition the variance in the predicted 24 h AFM1 reduction among model parameters.
2.5. Regulatory Benchmark And Calculation Of AFM1 Elimination
Regulatory compliance was assessed against the European Union maximum residue limit (MRL) for AFM1 in raw and heat‑treated milk of 0.05 µg/kg (approximately 0.05 µg/L, given milk density ≈1.03 kg/L)
. This is the strictest international standard; the Codex Alimentarius recommends a higher limit of 0.5 µg/L for some dairy products, but the EU MRL was used as the benchmark for this modelling exercise.The theoretical elimination of AFM1 was calculated by solving the first‑order differential equation:
where C is AFM1 concentration (µg/L), t is time (h), and keff (h⁻¹) is the effective removal rate constant. The analytical solution is:
For Model A (first‑order linear decay):
where k0 is the natural decay constant, α is the dose‑specific potency coefficient, and D is curcumin dose (mg/L).
For Model B (Michaelis–Menten saturation):
where Vmax is the maximum removal rate and Km is the half‑saturation constant.
Many physicochemical factors influence AFM1–curcumin–protein interactions, including temperature, pH, fat content, and mixing efficiency. In this in silico framework, these factors are not modelled explicitly. Instead, their combined effects are represented indirectly through the prior uncertainty distributions assigned to Vmax, Km, and C0 (Table 1), which were derived from literature on polyphenol–protein binding in dairy systems
, , . The model assumes constant temperature (4 °C, representing refrigerated storage) and homogeneous mixing; temperature dependence and fat partitioning are identified as key limitations requiring experimental characterisation.3. Main results
3.1. Kinetic Concentration Profiles
Fig. 1 presents mean kinetic trajectories over 24 hours under both models at each curcumin dose. In the untreated control (0 mg/L), AFM1 levels showed only minor spontaneous decline, consistent with the known chemical and thermal stability of this mycotoxin , . In curcumin-treated groups, Model A predicted a continuous, dose-proportional increase in removal efficacy up to 10 mg/L. In contrast, Model B revealed progressively diminishing returns, with a distinct performance plateau emerging between the 5 mg/L and 10 mg/L dose levels.
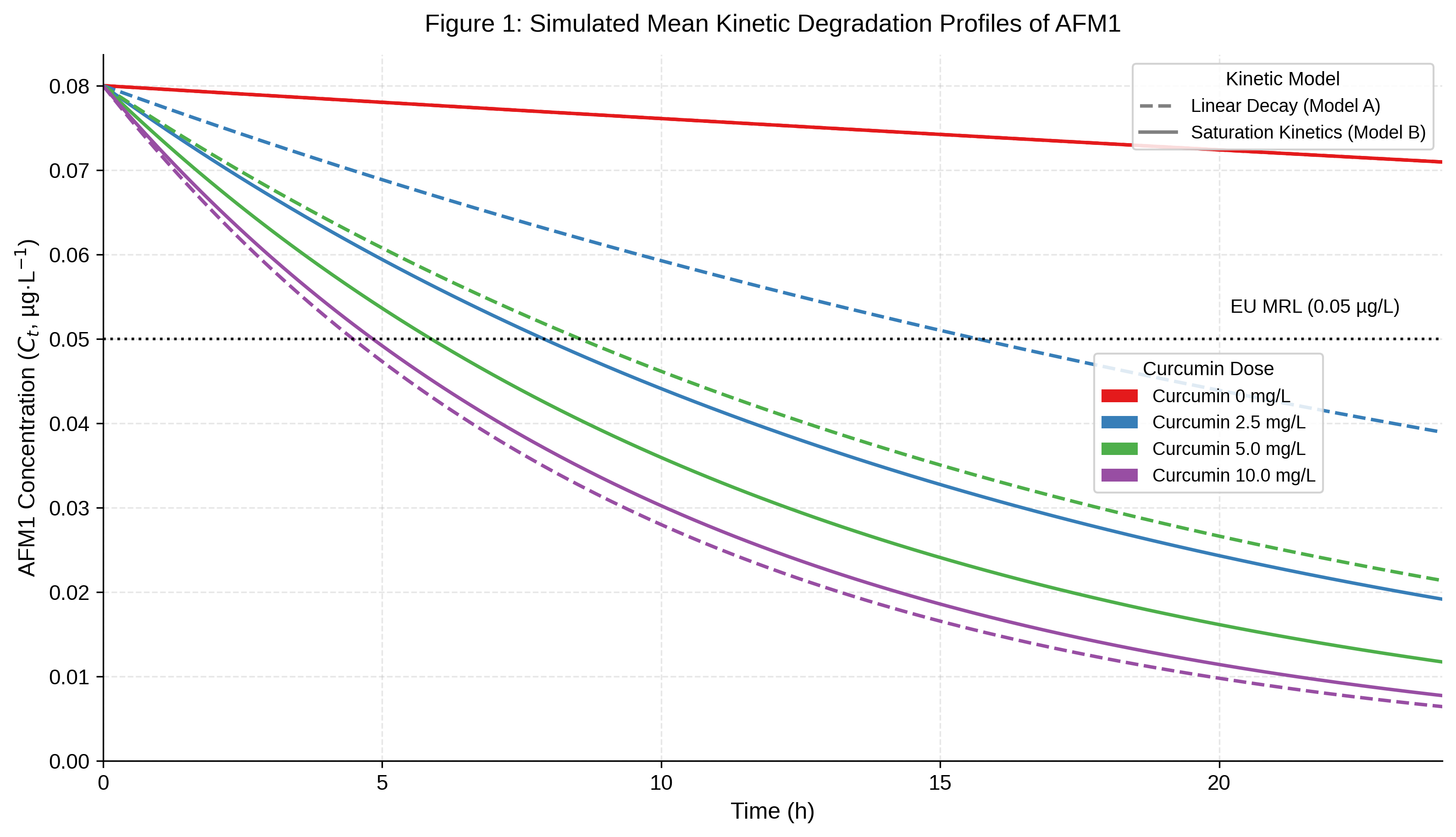
Mean kinetic degradation profiles of AFM1 in milk over 24 h under varying curcumin concentrations (0–10 mg/L):
dashed lines – Model A (first-order linear decay); solid lines – Model B (Michaelis–Menten saturation); red – 0 mg/L, blue – 2.5 mg/L, green – 5 mg/L, purple – 10 mg/L; dotted horizontal line – EU MRL (0.05 µg/kg)
At 10 mg/L, Model B predicted a residual AFM1 concentration approximately 20% higher than the corresponding linear model prediction. The saturation curve generated by Model B closely resembles binding isotherms reported in experimental studies of polyphenol–milk protein systems , , lending face validity to the saturation hypothesis. Future experimental designs must account for this matrix effect: performance data obtained in phosphate-buffered saline will underestimate the width of the predictive uncertainty distribution in whole milk and will not reveal the sub-saturation advantage of the Michaelis–Menten kinetics at lower doses.
3.2.1. Regulatory compliance probability
The distribution of final AFM1 concentrations at 24 h under 10 mg/L curcumin is shown in Fig. 2. Model A generated a narrow distribution centred near 0.007 µg/L, with 100% of Monte Carlo outcomes falling below the EU MRL. Model B produced a broader distribution centred near 0.008 µg/L, also achieving 100% compliance at this dose. At 2.5 mg/L, Model B attained 98% compliance versus 85% for Model A. At 5 mg/L, both models achieved compliance ≥ 99.6%. These figures represent unconditional probabilities across the full sampled C₀ distribution; approximately 7% of draws start already below the MRL at t = 0.
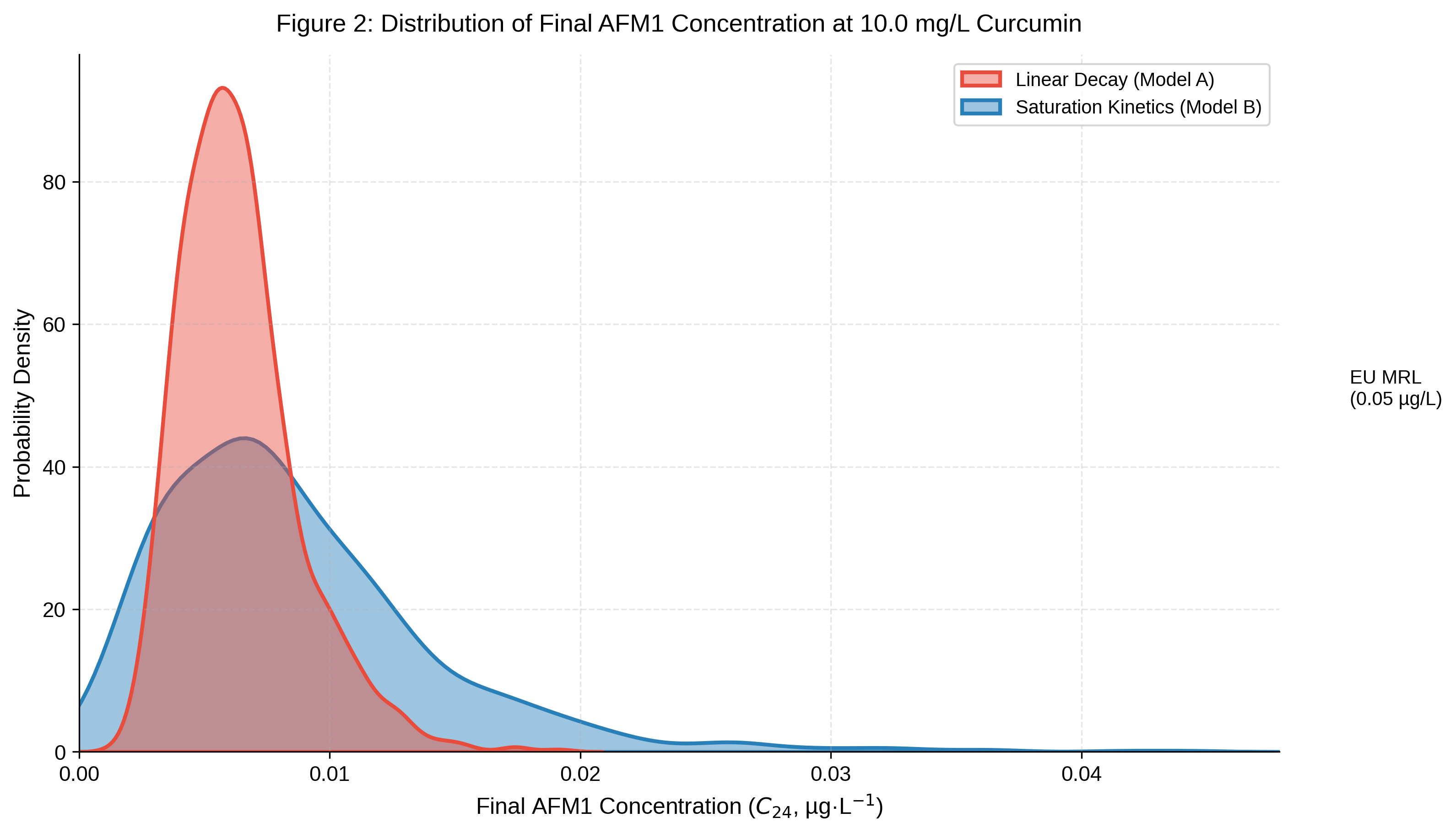
Kernel density distributions of final AFM1 concentration (C24 at 24 h under a curcumin dose of 10 mg/L, derived from 1000 Monte Carlo iterations:
red fill/dashed line – Model A (linear decay); blue fill/solid line – Model B (saturation kinetics); dotted vertical line – EU MRL (0.05 µg/kg)
First-order Sobol sensitivity indices for Model B at 10 mg/L are presented in Fig. 3. Vmax was the dominant source of predictive variance (Si = 0.697), contributing approximately 70% of total variance in AFM1 reduction at 24 h. The initial AFM1 concentration ranked second (Si = 0.160), followed by the half-saturation constant (Sᵢ = 0.135); k0 contributed negligible variance (Si = 0.000). The notably high contribution of C0 has a direct practical implication: tighter upstream control of feed-level aflatoxin B1, compressing the C0 distribution, would reduce model output uncertainty almost as effectively as experimental determination of Km. This hierarchy has direct experimental implications: uncertainty about Vmax, which represents the ceiling of curcumin efficacy determined by the degree of matrix interference, is the most critical unknown.
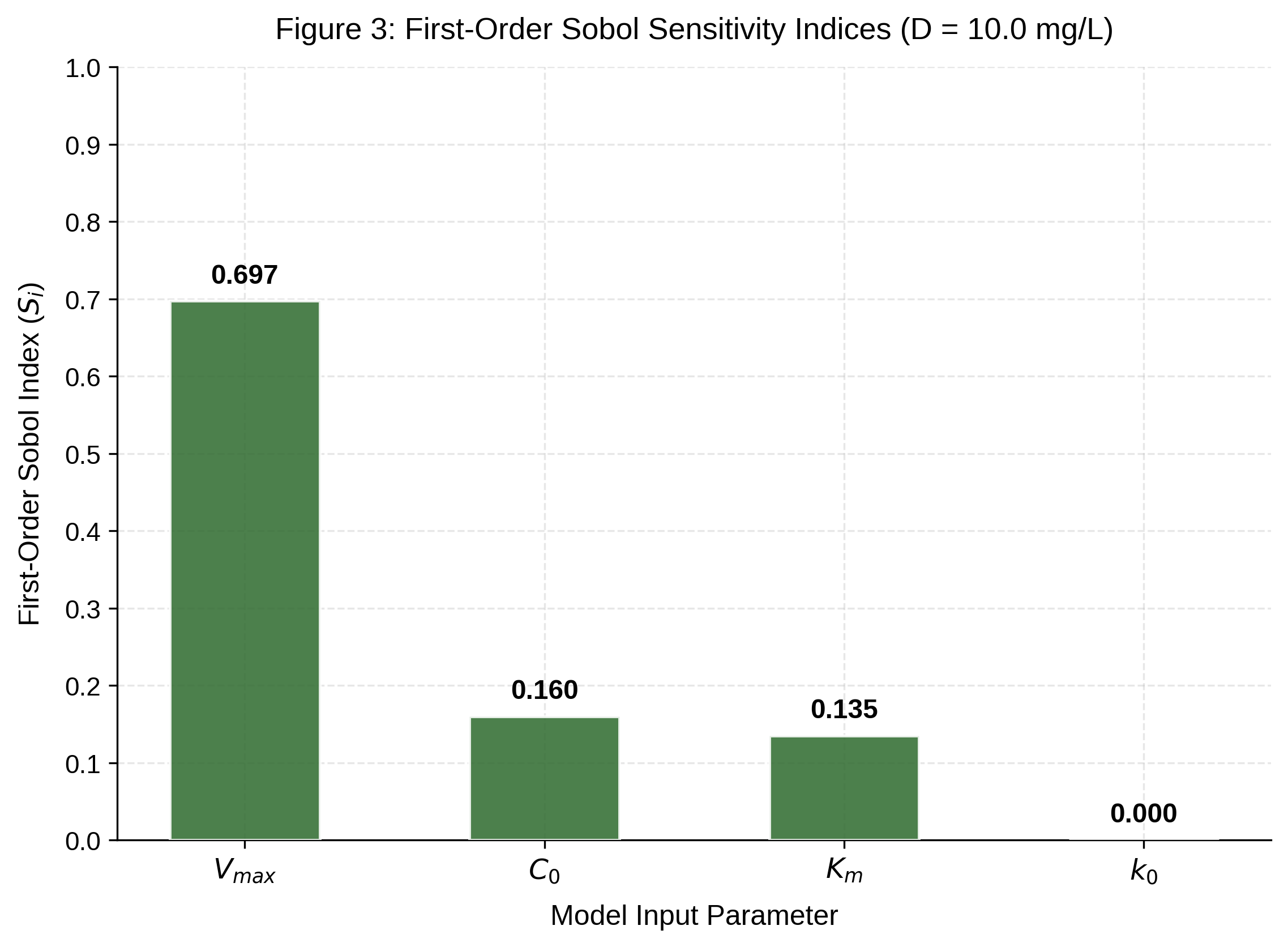
First-order Sobol sensitivity indices (Sᵢ) for the Michaelis–Menten saturation model (Model B) at 10 mg/L curcumin, showing the relative contribution of each input parameter to variance in predicted AFM1 reduction after 24 h
4. Discussion
The mechanistic basis for divergence between the two models is grounded in the physical chemistry of the milk matrix. Milk is a colloidal system rich in casein micelles and globular whey proteins, both of which are well-established binders of hydrophobic polyphenols , . Studies have demonstrated that curcumin forms stable multi-site complexes with micellar casein and whey protein fractions , , reducing the free fraction available for interaction with AFM1. Related investigations confirm that milk protein fractions strongly bind structurally related polyphenols and hydrophobic ligands , , , . In Model B, competitive protein sequestration is formalised within the Michaelis–Menten framework: as curcumin dose greatly exceeds Km, the effective removal rate kerr approaches the ceiling k0 + Vmax and ceases to respond proportionally to further dose increases. At 10 mg/L, curcumin represents a substantial molar excess relative to nanogram-level AFM1, so saturation is attributable not to AFM1 excess but to protein-mediated scavenging of curcumin, consistent with the broader polyphenol–protein competition literature , .
A notable counterintuitive result is that at low doses (2.5 mg/L), Model B predicts more rapid removal than Model A. This becomes clear when the two effective rate constants are compared: at D = 2.5 mg/L, the Michaelis–Menten effective rate is Vmax × D / (Km + D) = 0.12 × 2.5 / (3.0 + 2.5) ≈ 0.055 h⁻¹, whereas the linear effective rate is α × D = 0.010 × 2.5 = 0.025 h⁻¹. At sub-saturation doses where D ≪ Km, the Michaelis–Menten rate therefore exceeds the shallow linear slope defined by α; only once D ≫ Km does the linear model begin to outpace the saturation model. Buffer-based assay data would be expected to show linear, dose-proportional removal kinetics, providing false precision rather than simply overestimating mean removal when extrapolated to whole-milk systems .
The broader predictive distribution generated by Model B reflects the greater uncertainty inherent in Michaelis–Menten parameterisation and is consistent with published observations on the variability of polyphenol–protein binding across different milk compositions , . From a practical perspective, curcumin could serve as a primary or complementary hurdle for AFM1 mitigation; combining it with fermentative or adsorptive decontamination treatments , may further enhance reliability. More precise experimental determination of Vmax and Km under whole-milk conditions would substantially reduce predictive uncertainty. Advanced spectroscopic techniques, including fluorescence quenching, surface plasmon resonance, and ATR-FTIR, have been recommended for elucidating polyphenol–protein binding in dairy systems , , and their application to curcumin–casein and curcumin–AFM1 competition in full-fat versus skim milk would be of particular value , , .
This investigation carries several limitations. All kinetic parameters were drawn from analogical literature sources rather than direct experimental determination, as curcumin–AFM1 interaction constants in whole milk remain experimentally unavailable. The model assumes homogeneous mixing and does not account for fat globule partitioning of the lipophilic curcumin molecule, pH variation, or temperature-dependent effects on binding affinities. Potential degradation products of AFM1 are not considered. Future in vitro validation studies should determine binding isotherm constants in full-fat and skim milk separately (use spectroscopic or chromatographic approaches to quantify the free versus protein-bound curcumin fraction ); and consider mass balance modelling and molecular docking to refine mechanistic interpretation . Moreover, while curcumin is a permitted food additive (E100) and is well tolerated in many dairy applications, sensory trials in fluid milk would be required before industrial adoption. This does not diminish the value of curcumin as a decontaminant, but it does position it as most suitable for coloured or flavoured dairy products where such changes are acceptable or desirable.
5. Conclusion
A Monte Carlo simulation framework was applied to compare first-order linear and Michaelis–Menten saturation kinetic models for AFM1 reduction in milk by curcumin. The saturation model, which accounts for competitive sequestration of curcumin by milk matrix proteins, predicted a performance plateau at higher curcumin doses not captured by the linear model. At curcumin doses of 2.5 mg/L and above, both models predicted near-complete regulatory compliance with the EU MRL of 0.05 µg/kg. At 2.5 mg/L, Model B achieved notably higher compliance (98%) than Model A (85%), reflecting more favourable sub-saturation kinetics. The saturation model generated a substantially broader predictive distribution at all doses, indicating that parametric uncertainty, rather than mean efficacy, is the key limitation of this framework.
Sobol sensitivity analysis identified Vmax (Si = 0.697) as the overwhelmingly dominant source of predictive uncertainty, followed by C0 (Si = 0.160) and Km (Si = 0.135); k₀ contributed negligible variance (Si = 0.000). Experimental determination of Vmax and Km in whole milk remains the most important near-term research priority. The probabilistic framework presented provides a practical, uncertainty-aware basis for designing targeted in vitro experiments and for evaluating curcumin as a component of multi-hurdle AFM1 mitigation strategies in dairy systems.