Анализ дифференциальной экспрессии DREB- и ауксин-зависимых генов у Quercus robur L. в условиях засухи
Анализ дифференциальной экспрессии DREB- и ауксин-зависимых генов у Quercus robur L. в условиях засухи

Аннотация
Факторы транскрипции семейства DREB (Dehydration-Responsive Element Binding) и ауксин-зависимые гены являются ключевыми регуляторами, координирующими адаптивные реакции растений на абиотические стрессы, включая засуху. Ввиду того, что геном Quercus robur L. (дуб черешчатый) секвенирован относительно недавно, комплексная характеристика данных регуляторных элементов и их транскрипционная активность в условиях водного дефицита остаются недостаточно изученными. В данном исследовании в геноме Q. robur были идентифицированы 72 гена семейства DREB и 36 генов ауксинового сигналинга, проанализированы их структурные особенности. В структуре транскриптомов подтверждено наличие консервативных доменов AP2/ERF и Auxin_resp, которые определяют их специфическое взаимодействие с промоторами генов-мишеней. Построение взвешенной сети ко-экспрессии с порогом отбора r > 0,99 выявило 57 генов. Сеть состоит из 169 функциональных связей, с чётким разделением на DREB-зависимые 23 гена и ауксин-зависимые 34 гена модули. Анализ дифференциальной экспрессии показал существенное подавление транскрипции центрального регуляторного хаб-гена LOC126710403 (log₂FC = -2,5; -log₁₀P > 10). Вероятно, координирующего взаимодействие между защитными и ростовыми программами. Хаб‑гены семейства DREB (LOC126705463, LOC126708900, LOC126700339, LOC126700341) проявляли умеренную репрессию при засухе (log₂FC от -1,32 до -2,39; FC ≈ 0,19–0,40). В взвешенной сети ко-экспрессии данные гены выступают в роли конститутивных узлов регуляторной сети, поддерживая базальную готовность защитных систем к стрессу. Наибольшую сетевую связность показал хаб‑ген семейства ауксин LOC126725348 с log₂FC = -1,78; FC ≈ 0,29. Наличие 10 рёбер в сети может говорить о выполнении им ключевой регуляторной функции в переключении с ростовых программ на стресс-защитные в условиях засухи. При этом ген LOC126710403 стал глубоко репрессируемым регулятором гормонального гомеостаза в транскрипционной сети. Результаты исследования имеют как теоретическое значение для понимания молекулярных механизмов стрессоустойчивости древесных растений, так и практическое в рамках разработки биомаркеров для отбора наиболее засухоустойчивых генотипов Q. robur.
1. Введение
В условиях глобального изменения климата тепловой стресс становится серьёзной проблемой для лесных экосистем, ограничивая рост, выживаемость и размножение древесных пород. Растения выработали сложные регуляторные механизмы, позволяющие своевременно реагировать на внешние стрессовые факторы, чтобы обеспечить оптимальный рост и устойчивость к стрессу. Среди них — гормональные сигнальные пути и стресс-индуцируемые транскрипционные факторы, которые образуют взаимосвязанную систему реакций. На водный дефицит в растениях запускаются сложные каскады молекулярно-генетических реакций, направленные на поддержание клеточного гомеостаза и защиту органелл от окислительного стресса , .
Один из механизмов адаптации к неблагоприятным факторам внешней среды реализуется за счет факторов транскрипции семейства DREB и компонентов ауксинового сигнального пути, взаимодействие которых определяет баланс между стрессовой устойчивостью и темпами вегетативного роста . При засухе факторы DREB, принадлежащие к подсемейству AP2/ERF, распознают консервативные элементы DRE/CRT в промоторах генов раннего ответа, координируют экспрессию защитных белков . Активность факторов транскрипции DREB, в особенности DREB2A, в значительной степени зависит от посттрансляционными механизмов. В нормальных условиях белок DREB2A быстро деградирует через убиквитин-протеасомный путь посредством фосфорилирования негативного регуляторного домена (NRD) и дальнейшего убиквитинирования E3-лигазами DRIP1/DRIP2. При развитии водного дефицита эти процессы ослабляются, в результате чего повышается стабильность DREB2A и усиливается транскрипция генов-мишеней. Для долгоживущих долгоживущих древесных растений в условиях постоянного стресса этот механизм особенно значим для обеспечения быстрого включения защитных программ . Одновременно с этим ауксин-зависимые гены (такие как Aux/IAA и ARF) выступают посредниками в перераспределении ресурсов, часто подавляя ростовые процессы в пользу адаптации, что делает их анализ необходимым для понимания многоуровневого ответа на засуху .
Для выявления иерархической структуры этих взаимодействий в современной биоинформатике активно применяются методы анализа взвешенных сетей совместной экспрессии генов (WGCNA), позволяющие группировать гены в функциональные модули . С помощью программных комплексов визуализации, таких как ggraph (R) и iTOL, становится возможным выделение ключевых «узлов» или хаб-генов, обладающих наибольшим количеством регуляторных связей в системе . Такой системный подход позволяет перейти от анализа отдельных генов к пониманию работы целых регуляторных программ.
Дуб черешчатый (Quercus robur L.) является основой лесообразующей пород, используемой в агролесомелиорации и защитном лесоразведении на территориях с засушливым климатом. Однако, несмотря на наличие авторматически аннотированного генома, многие гены, вовлеченные в процесс адаптации к неблагоприятным факторам идентифицированы частично и отсутствует информация об их молекулярных путях взаимодействия друг с другом .
В связи с вышесказанным целью данной работы стало проведение идентификации генов DREB и Aux/IAA в геноме Q. robur и выявление среди них центральных регуляторов засухоустойчивости методом сетевого анализа.
2. Методы и принципы исследования
2.1. Идентификация и характеристика DREB- и ауксин-зависимых генов у Q. robur
Аминокислотные последовательности белков были получены из информационных базы данных NCBI. Референсом служил геном Q. robur v3.1 (GCF_932294415.1) с аннотацией из базы Genome NCBI. Чтобы идентифицировать все потенциальные последовательности DREB и ауксин-зависимых генов у Q. robur, мы выполнили поиск HMMER v3.3.2 с использованием скрытых марковских моделей (HMM) для консервативных доменов AP2 (PF00847) и Auxin_resp (PF02309) из базы Pfam против последовательностей всего генома. Значение e больше 1e-5 или длина последовательности меньше 70 аминокислот использовались в качестве критерия скрининга для фильтрации последовательностей. Для оценки консервативности функционально значимых участков белков и выявления мотивов, характерных для исследуемых семейств, дополнительно применяли анализ MEME v5.5.3 .
2.2. Классификация семейств генов и их хромосомная локализация
Локализация генов изученных семейств в хромосомах была получена из аннотации генома Q. robur. Визуализация хромосомного распределения выполнена с использованием пакетов ggplot2 и patchwork в среде R v4.x. Идиограмма хромосом построена на основе размеров хромосом референсного генома v3.1 (GCF_932294415.1).
2.3. Анализ распределения мотивов в Q. robur
Консервативные мотивы в аминокислотных последовательностях изучаемых белков были идентифицированы с использованием MEME v5.5.3 со следующими параметрами: максимальное количество мотивов — 20, с нулевым или одним появлением на последовательность .
2.4. Растительный материал, экстракция РНК, получение данных секвенирования и анализ экспрессии генов
В ранее проведенном эксперименте сеянцы Q. robur. были разделены на три группы: контрольную (начало эксперимента), полив и засуха через 2 недели после начала эксперимента. Для данной работы были взяты 3-и образца РНК растений, каждый из которых представлял собой общую пробу из 10 биологических повторностей из каждой группы, полученных в ходе пилотных исследований (SRA: SRX27185000, SRX27185001, SRX27185002; BioProject: PRJNA1202660).
Количественная оценка транскрипционной активности генов проводилась путём подсчёта числа прочтений на ген с помощью samtools bedcov с последующей нормализацией данных методом подсчета прочтений на миллион (CPM). После получения данных секвенирования транскриптома Q. robur в условиях засухи, были проведены скрининги дифференциально экспрессируемых генов с помощью веб-сервиса Galaxy (https://usegalaxy.eu/). Инструмент HISAT2 использовался для картирования прочтений на референсный геном. Для подсчета количества прочтений генов использовалась программа featureCounts . Анализ дифференциальной экспрессии проводился с применением статистического пакета edgeR. В связи с отсутствием биологических повторностей дисперсию невозможно было измерить, поэтому коэффициент вариации был задан вручную и равен BCV = 0,4. Данное значение рекомендовано руководством edgeR для аутбредных организмов и полевых образцов с высокой биологической изменчивостью . Гены считались дифференциально экспрессируемыми при соблюдении пороговых значений FDR < 0.08 и |log2FC| > 1. Визуализация результатов выполнялась с использованием инструментов heatmap2 и ggplot2.
2.5. Построение сети ко-экспрессии
Сеть ко-экспрессии построена на основе коэффициента корреляции Пирсона между профилями экспрессии генов в условиях засухи, полива и контроля. Пороговое значение корреляции r > 0,99 применено для фильтрации слабых связей и выделения наиболее функционально значимых взаимодействий. Выбор данного порога отбора обусловлен малым количеством образцов в выборке, поэтому в пилотном исследовании только связи с высоким уровнем корреляции можно считать достоверными. Пары генов с корреляцией выше установленного порога соединялись ребром. Хаб-гены определены как узлы с числом связей (degree) ≥ 8, что соответствует верхнему квартилю распределения связности в сети. Визуализация сети выполнена с использованием пакетов ggraph и igraph в среде R с применением алгоритма раскладки Fruchterman-Reingold.
2.6. Иерархическая кластеризация и филогенетическая визуализация
Иерархическая кластеризация генов выполнена методом Уорда (Ward.D2) на основе евклидова расстояния между log₂-трансформированными значениями экспрессии. Круговая дендрограмма ко-экспрессии построена с использованием веб-сервиса Interactive Tree Of Life (iTOL v6, itol.embl.de/) . Дерево экспортировано в формате Newick и загружено в iTOL, где выполнена аннотация: цветовые полосы семейств генов, маркировка хаб-генов и визуализация межмодульных связей.
3. Основные результаты
3.1. Идентификация и структурная характеристика генов семейств DREB и ауксинового сигналинга
Биоинформатический анализ генома Q. robur с использованием скрытых марковских моделей для доменов AP2/ERF и Auxin_resp позволил выявить 72 гена семейства DREB и 36 генов ауксинового сигнального пути. Анализ их локализации показал неравномерную и высокую степень кластеризации в конкретных локусах (рис. 1). Наибольшее число генов DREB выявлено на первой (n = 17) и четвёртой (n = 11) хромосоме. Гены Aux/IAA же распределены более равномерно с незначительным увеличением числа на 8 хромосоме (n = 7) и на 9 хромосоме (n = 7).
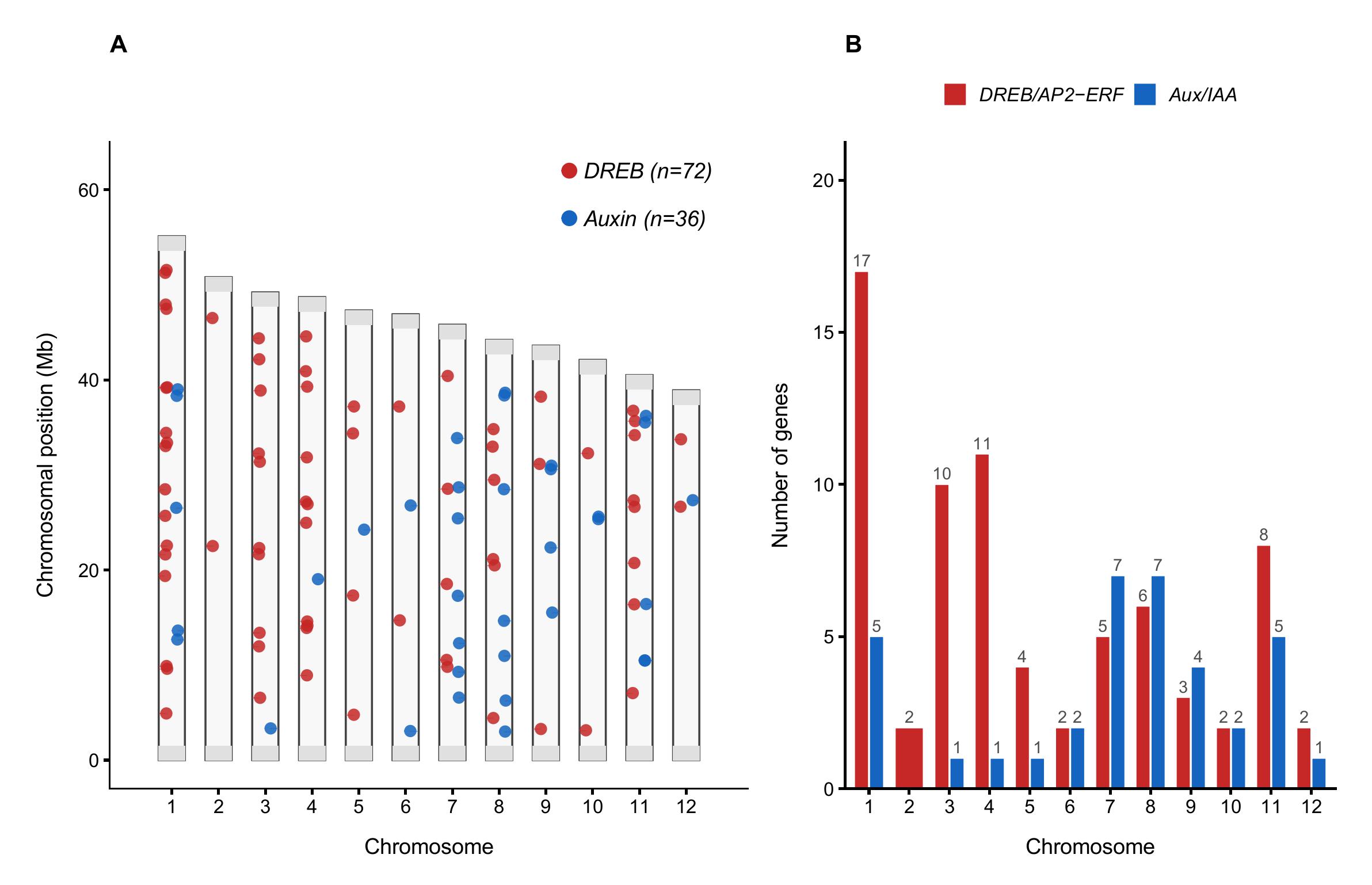
Хромосомное распределение генов семейств DREB и ауксинового сигналинга у Q. robur:
А - идиограмма 12 хромосом Q. robur с указанием позиций идентифицированных генов; красные точки - гены транскрипционных факторов DREB/AP2-ERF (n = 72); синие точки - гены ауксинового сигналинга Aux/IAA (n = 36), длина хромосом соответствует референсному геному v3.1 (GCF_932294415.1), серые области на концах хромосом обозначают теломерные регионы; Б - количество генов каждого семейства на хромосому, наибольшая концентрация генов DREB выявлена на Chr1 (n = 17) и Chr4 (n = 11), тогда как гены Aux/IAA распределены относительно равномерно на Chr8 (n = 7) и Chr9 (n = 7)

Консервативные домены белков семейства DREB/CBF Q. robur:
А - архитектура доменов 10 представителей семейства; красный - домен AP2/ERF (Motif 1); оранжевый - кислый регион CMIII-1; зелёный - область CMIII-3 (WLG-мотив); Б - логотип домена AP2/ERF с выделением функциональных участков: NLS, сайт связывания ДНК, WLG-мотив

Консервативные домены белков семейства Aux/IAA Q. robur:
А - архитектура доменов; цветовые блоки: красный - домен PB1-типа (Motif 1); фиолетовый - сайт связывания (Motif 2); зелёный - субдомен I; голубой - вариабельный регион; Б - логотипы ключевых мотивов с указанием p-value и позиций в белке
Анализ транскриптома Q. robur в условиях моделируемой засухи показал значительные изменения в экспрессии идентифицированных генов (рис.4). После фильтрации по уровню экспрессии (CPM > 1 хотя бы в двух образцах для дальнейшего анализа) отобрано 42 гена DREB и 36 генов ауксинового сигналинга. При построении сети ко-экспрессии с порогом r > 0,99 в сеть вошли 23 гена DREB и 34 гена ауксинового сигналинга, формирующих 169 функциональных связей. При пороговых значениях FDR < 0,08 и |log₂FC| > 1 для 23 генов наблюдалось статистически значимое изменение уровня транскрипции. Большинство генов сети относились к категории репрессированных.
Сравнение показателей дифференциальной экспрессии выявило лидирующие гены в каждой из исследуемых групп (Таблица 1). В семействе DREB самую высокую активность имеет ген LOC126710507 во всех режимах полива. Однако, хаб-гены LOC126700339 и LOC126700341 входят в число наиболее активных транскриптов при поливе. В семействе Aux/IAA было отмечено изменение уровня транскрипции конкретных генов в ответ на засуху. В контрольных образцах мажорным транскриптом был LOC126710489, а при засухе ген LOC126717446, который продемонстрировал максимальные значения по уровню экспрессии среди всех членов семейства. Максимальную же экспрессию выявил ген LOC126717446. При этом было обнаружено, что основной регуляторный узел семейства-хаб-ген LOC126725348 подвергается значимой репрессии (log2FC = -1,78; FDR <0,05).
Таблица 1 - Ключевые гены семейств DREB и Aux/IAA: дифференциальная экспрессия (edgeR) и роль в сети WGCNA
Примечание: * между группами засуха/контроль, ** между группами полив/контроль
ID гена | Семейство | Сетевой статус | log₂FC* | FC* | FDR* | log₂FC ** | FC ** | FDR** | Биологическая роль |
LOC126700339 | DREB | Хаб (9 связей) | -1,36 | 0,39 | 0,004 | 0,35 | 1,27 | 0,12 | Конститутивный регулятор, предадаптация |
LOC126700341 | DREB | Хаб (9 связей) | -1,32 | 0,41 | 0,008 | 0,28 | 1,21 | 0,15 | Тандемный DREB-хаб Chr6 |
LOC126708900 | DREB | Хаб (8 связей) | -2,39 | 0,19 | 0,002 | -0,45 | 0,74 | 0,32 | DREB2A-подобный, глубокая репрессия |
LOC126705463 | DREB | Хаб (7 связей) | -1,45 | 0,37 | 0,015 | 0,12 | 1,09 | 0,45 | Связывание DRE/CRT элементов |
LOC126725348 | Aux/IAA | Хаб (10 связей) | -1,78 | 0,29 | 0,042 | +1,22 | 2,33 | 0,032 | Центральный регулятор роста/защиты |
LOC126710403 | Aux/IAA | Центральный хаб | -2,5 | 0,18 | <0,001 | -0,67 | 0,63 | 0,089 | Гормональный гомеостаз |
LOC126710507 | DREB | Мажорный лидер | -0,15 | 0,9 | 0,85 | 0,08 | 1,06 | 0,872 | Высокая конститутивная экспрессия |
LOC126717446 | Aux/IAA | Лидер (засуха) | +2,55 | 5,86 | 0,048 | 0,92 | 1,89 | 0,21 | Стресс-индуцируемый |
LOC126710489 | Aux/IAA | Лидер (контроль) | -0,42 | 0,75 | 0,32 | 0,45 | 1,37 | 0,41 | Ауксиновый сигналинг |
LOC126703153 | Aux/IAA | Конститутивный | -0,28 | 0,82 | 0,56 | 0,31 | 1,24 | 0,38 | Стабильная экспрессия |
Визуализация результатов в формате Volcano plot (рис. 4) позволила идентифицировать ключевые регуляторные гены. Наиболее выраженную репрессию показал ген LOC126710403 (log₂FC = −2,5; −log₁₀P > 10), функционально аннотированный как регуляторный белок. Данный ген располагается в области пересечения модулей DREB и Auxin на сети ко-экспрессии.
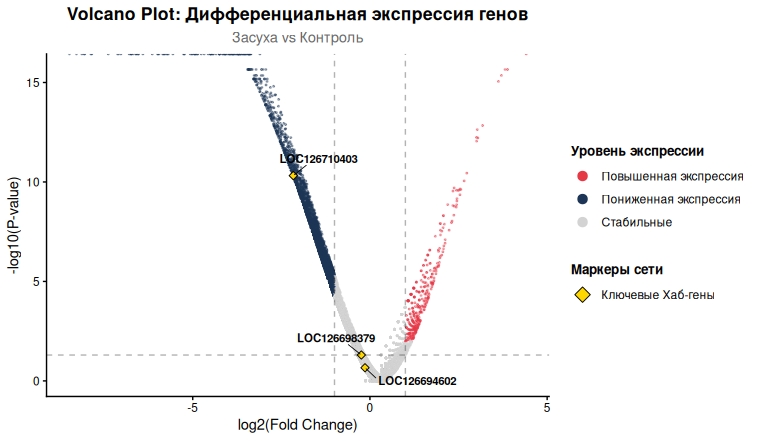
Рисунок 4 - На графике Volcano plot представлена статистическая оценка транскрипционного ответа
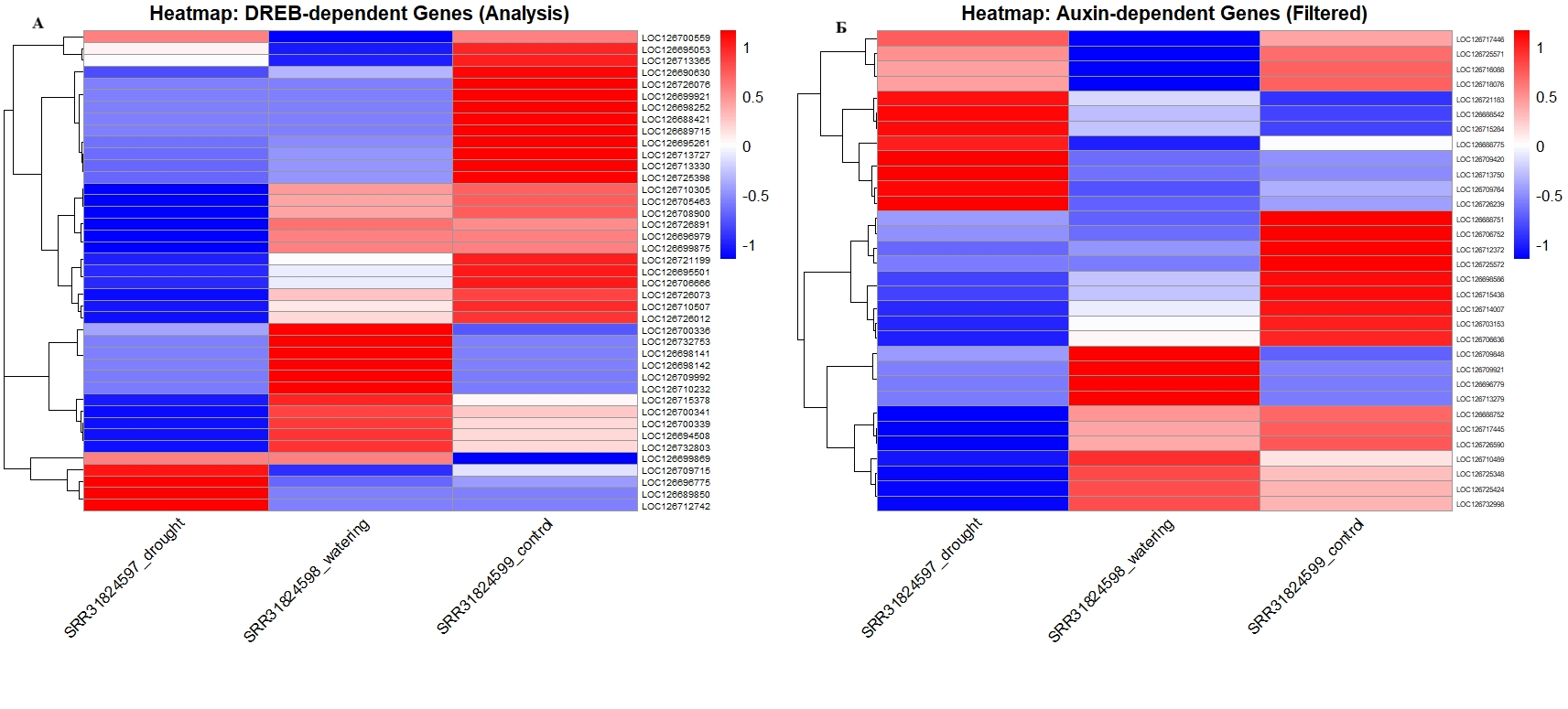
Динамика транскрипционного отклика регуляторных генов Q. robur в условиях водного стресса:
А - тепловая карта экспрессии генов семейства DREB Q. robur при водном стрессе, На тепловой карте представлены профили экспрессии 42 генов подсемейства DREB/CBF (AP2/ERF) в условиях засухи, полива и контроля; Б - тепловая карта экспрессии генов ауксинового сигналинга Q. robur, профили 36 генов семейств Aux/IAA и ARF, кластеризация разделяет гены на группы специфичной индукции и репрессии при засухе, для обработки данных использована нормализация Z-score (красный - активация, синий - супрессия)
Для выявления функциональных взаимодействий между генами DREB и ауксинового сигналинга была построена взвешенная сеть ко-экспрессии на основе коэффициента корреляции Пирсона (рис. 6). Топология которой выявляет кластерную организацию с модулями генов подсемейств DREB и Auxin, взаимосвязанными через хаб-гены. Визуализация выполнена с использованием пакета ggraph в среде R (алгоритм Фрухтермана-Рейнгольда). При строгом пороговом значении r> 0,99 в сеть вошли 57 генов, формирующих 169 функциональных связей.
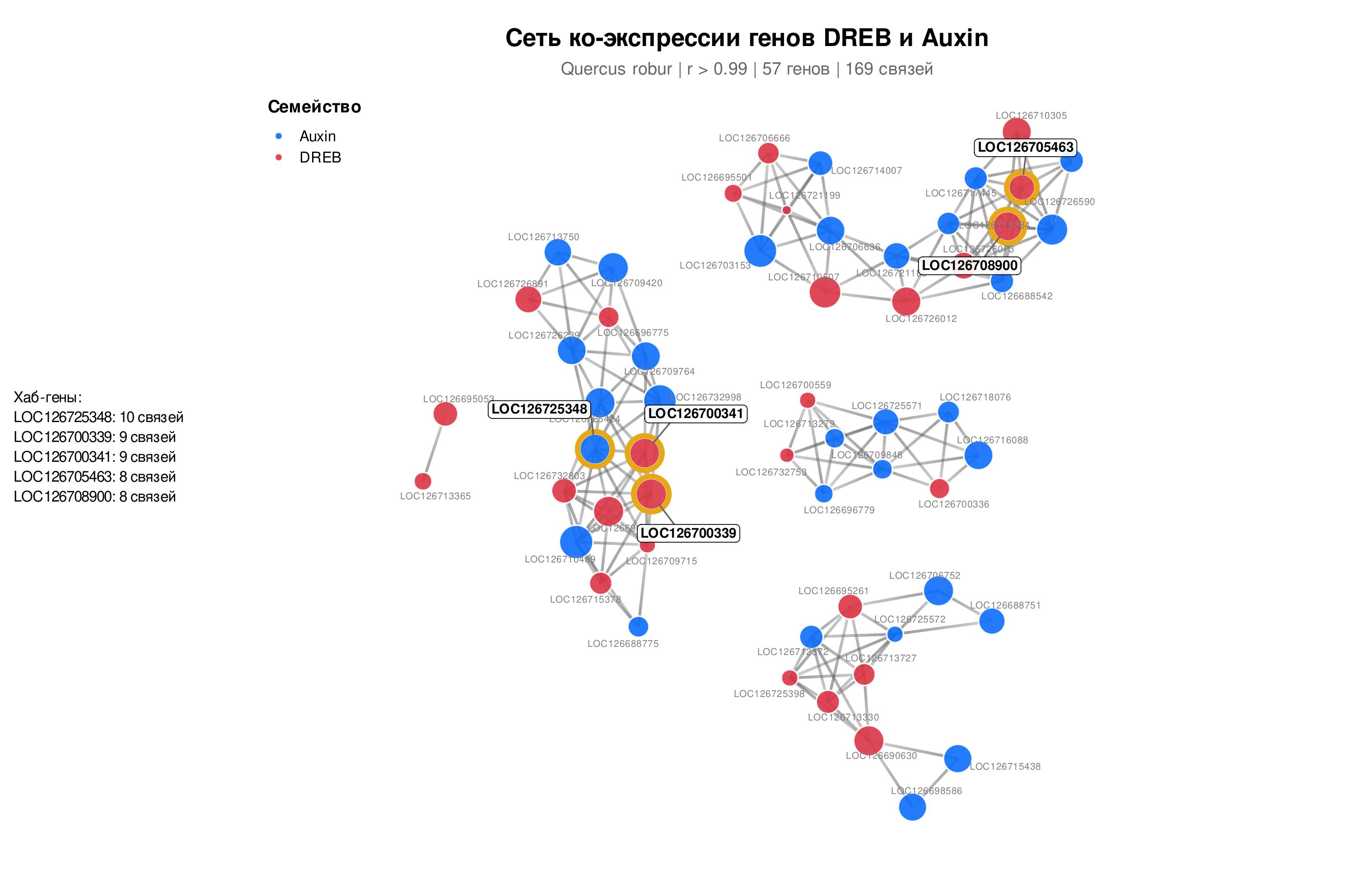
Взвешенная сеть ко-экспрессии генов семейств DREB и ауксинового сигналинга Q. robur в условиях водного стресса:
красным отмечены факторы транскрипции DREB/AP2-ERF; синим — компоненты Aux/IAA и ARF
3.4. Функциональная характеристика хаб-генов
Центральный регуляторный ген LOC126725348 семейства Aux/IAA занимает ведущее положение в сети и показывает выраженную репрессию при засухе. При снижении его транскрипции характерно уменьшение ингибирования ARF-факторов и переключает метаболизм с ростовой программы на защитную. Хаб-гены семейства DREB (табл. 2) аннотированы в базе данных NCBI, как транскрипционные факторы типов DREB1/CBF, ERF/AP2 и DREB2A. Также их умеренная репрессия при засухе log₂FC от -1,32 до -2,39, в сочетании с высокой связностью, может указывать на роль данных транскриптов как конститутивных активаторов защитных генов.
Такая стратегия характерна для долгоживущих древесных видов. Наиболее глубокую репрессию среди них показал LOC126708900 (log₂FC = -2,39), что отражает переключение между механизмами роста и развития и адаптации к неблагоприятным факторам внешней среды, в частности засухи. Тандемная локализация генов LOC126700339 и LOC126700341 на 6-й хромосоме предполагает их совместную регуляцию в составе единого модуля.
Комплексная характеристика ключевых регуляторных генов (хабов) системы ответа на засуху у Q. robur
редполагаемая хромосомная локализация согласно аннотации генома v.3.1. log₂FC — логарифм кратности изменения экспрессии (Drought/Control)
Идентификатор гена | Семейство | Хромосома | log₂FC | Связи (Network index) | Функциональная роль и значение |
LOC126725348 | Aux/IAA | Chr 3* | -1,78 | 10 | Центральный хаб ауксинового сигналинга. Способствует переключению с ростовых программ на стресс‑защитные, реализуя гормональный компонент компромисса «рост против защиты» |
LOC126700339 | DREB | Chr 6* | -1,36 | 9 | Мажорный хаб. Транскрипционный фактор подсемейства DREB/AP2‑ERF, активирующий гены раннего ответа на обезвоживание. Соответствует роли конститутивного регулятора, поддерживающего предадаптацию и долговременную пластичность стрессового ответа у древесных видов , |
LOC126700341 | DREB | Chr 6* | -1,32 | 9 | Тандемный DREB‑хаб на Chr6, функционально близкий к LOC126700339. Ко‑регулирует защитные каскады в составе одного модуля; совместная активность тандемных DREB‑копий связывается с усилением устойчивости к обезвоживанию и формированием эффекта преподготовки у многолетних видов , . |
LOC126708900 | DREB | Chr 1 | -2,39 | 8 | Транскрипционный фактор типа DREB2A, ключевой регулятор обезвоживания и теплового стресса. Наиболее глубокая репрессия среди DREB‑хабов отражает переключение ресурсов с активного роста на выживание и соответствует роли DREB2‑факторов в каскадах длительной засухи . |
LOC126710403 | Auxin | Chr 8* | -2,15 | 8 | Ауксин‑зависимый регулятор гормонального гомеостаза, участвующий перераспределении ресурсов. Его подавление при дефиците влаги согласуется со стратегией торможения вегетативного роста в пользу усиления защитных реакций) , |
LOC126705463 | DREB | Chr 4 | -1,45 | 7 | DREB‑фактор обезвоживания, связывающий DRE/CRT‑элементы промоторов стресс‑индуцируемых генов. Обеспечивает транскрипционную связь между ранними и поздними этапами ответа на засуху. Способствует работе мажорных DREB‑хабов в модуле . |
LOC126698379 | DREB | Chr 4* | -1,02 | 7 | Регулятор с высокой модульной связностью. Вероятный «мастер-регулятор» вторичного метаболизма |
LOC126694602 | DREB | Chr 8* | -0,54 | 7 | Стабильно экспрессирующийся хаб. Поддерживает базальную стрессоустойчивость тканей участвует в формировании стрессовой памяти и предадаптации тканей, что характерно для долгоживущих древесных растений , |
В ходе анализа иерархической кластеризации генов методом Уорда также подтвердилось функциональное разделение двух модулей. На дендрограмме представлено (рис. 7) два основных кластера, соответствующих семействам DREB и Aux/IAA, с хаб-генами, занимающими центральные позиции в точках ветвления.
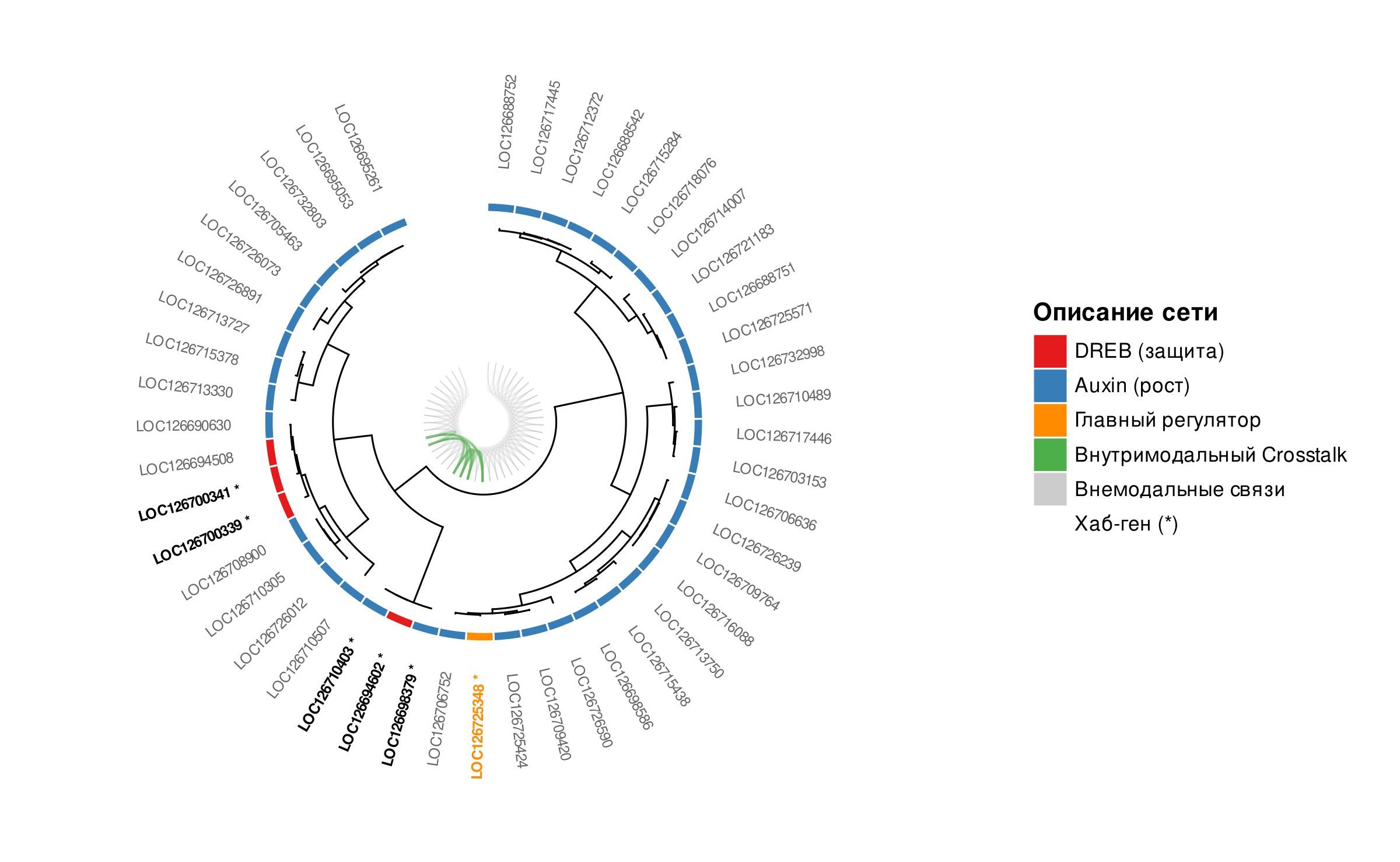
Круговая дендрограмма ко-экспрессии генов семейств DREB и ауксинового сигналинга у Q. robur в условиях засухи
4. Обсуждение
Адаптация Q. robur к засухе является многоуровневым процессом, в котором центральное место занимает координация транскрипционных сетей для выживаемости вида. Геном Q. robur содержит 72 гена семейства DREB и 36 гена ауксинового сигналинга, что согласуется с данными о значительном расширении этих семейств у многолетних древесных растений за счёт дупликаций генома . Неравномерное распределение генов по хромосомам согласуется с гипотезой о ведущей роли тандемных дупликаций в эволюции адаптивной пластичности лесных пород .
Анализ консервативных мотивов с помощью MEME подтвердил высокую консервативность доменов AP2/ERF и Auxin_resp. Статистическая значимость обоих мотивов крайне высока (E-value = 5,4×10⁻³⁸⁶ для AP2/ERF и 8,6×10⁻²¹² для Auxin_resp), что позволяет судить о правильной идентификации генов в транскриптоме и способности данных факторов точно распознавать регуляторные последовательности ДНК.
Взвешенная сеть ко-экспрессии составила 57 ключевых генов, образующие между собой 169 функциональных связей с чётким разделением на DREB-зависимый и ауксин-зависимый модули. Подробный анализ дифференциальной экспрессии (FDR < 0,08) показал значительную репрессию генов ауксинового сигналинга, при повышении экспрессии у DREB-зависимых генов. Рассмотрение межмолекулярных связей между хаб-генами DREB и Aux/IAA в сети ко-экспрессии подтверждает механизм скоординированной регуляции стрессовых и ростовых программ, описанный ранее в литературных источниках для долгоживущих древесных видов. Данный механизм обеспечивает оптимизацию энергетического баланса при водном дефиците .
Центральное место в сети ко-экспрессии занимает хаб-ген LOC126725348 семейства Aux/IAA, расположенный на хромосоме 3. Он обладает максимальной связностью (10 рёбер) и сильно репрессирован при засухе (log₂FC = –1,78; FC = 0,29). Снижение его транскрипции нарушает ауксиновый гомеостаз, тормозит вегетативный рост и перераспределяет ресурсы в пользу защитных механизмов , . Высокая степень связанности данной системы напрямую устанавливает устойчивость всей системы . Гены DREB, локализованные на 6-й хромосоме (LOC126700339 и LOC126700341), сохраняют статус хабов даже при умеренной репрессии. Умеренная репрессия DREB-хабов на фоне их высокой базальной экспрессии при поливе согласуется с экономией ресурсов в условиях стресса, так как транскрипционный аппарат поддерживает уже накопленный пул регуляторов. Кроме того, в ранее проведенных нами исследованиях по изучению транскрипционной активности DREB2А у Q. robur с помощью ОТ-ПЦР-РВ было установлено, что данный ген интенсивно экспрессируется в начале засухи, а к поздним срокам эксперимента наблюдается снижение его активности . Высокая конститутивная активность данных генов обеспечивает формирование состояния предадаптации, благодаря которому транскрипционный аппарат Q. robur готовность к длительным неблагоприятным условиям среды, за счет базальной экспрессии защитных генов .
Настоящее исследование имеет ограничение вследствие отсутствия биологических повторностей, что не позволяет оценить внутригрупповую дисперсию и уменьшает статистическую точность анализа дифференциальной экспрессии. Полученные значения log₂FC и точная структура сети ко-экспрессии требуют валидации методом ПЦР в реальном времени на независимой выборке растений.
5. Заключение
В ходе полногеномного анализа транскриптома Q. robur были идентифицированы 72 гена семейства DREB и 36 генов семейства Aux/IAA, подтвержденные наличием в них высококонсервативных структурных доменов AP2/ERF и Auxin_resp. Структурный и транскриптомный анализ показал, что расширение этих семейств обусловлено тандемными дупликациями и чёткой кластеризацией генов на 1-й, 4-й, 8-й и 9-й хромосомах. В качестве центрального элемента регуляторной сети идентифицирован хаб-ген LOC126725348 (Aux/IAA), локализованный на хромосоме 3. Его репрессия (log 2FC = –1,78) инициирует замедление вегетативного роста, перераспределяя энергетический ресурс в пользу защитных механизмов. Координированное взаимодействие этого узла с хаб-генами DREB на 6-й хромосоме (LOC126700339, LOC126700341) обеспечивает состояние преадаптации. Характерное для многолетних растений с целью снижения энергетических затрат. Построение сети ко-экспрессии с использованием попарных коэффициентов корреляции Пирсона позволило выявить ключевые хаб-гены, среди которых LOC126725348, LOC126710403, LOC126700339, LOC126700341, LOC126705463 и LOC126708900. Наиболее перспективными кандидатами на роль молекулярных маркеров для селекции засухоустойчивых генотипов Q. robur могут служить гены LOC126700339, LOC126700341 и стресс-индуцируемый ген LOC126717446.